Research Interests
The luteus group of Mimulus (black branches on the phylogeny, below) contains five monkeyflower species. All are native to central Chile; their ranges are shown on the map below. They form a small, monophyletic group characterized by a sudden burst of floral diversification.
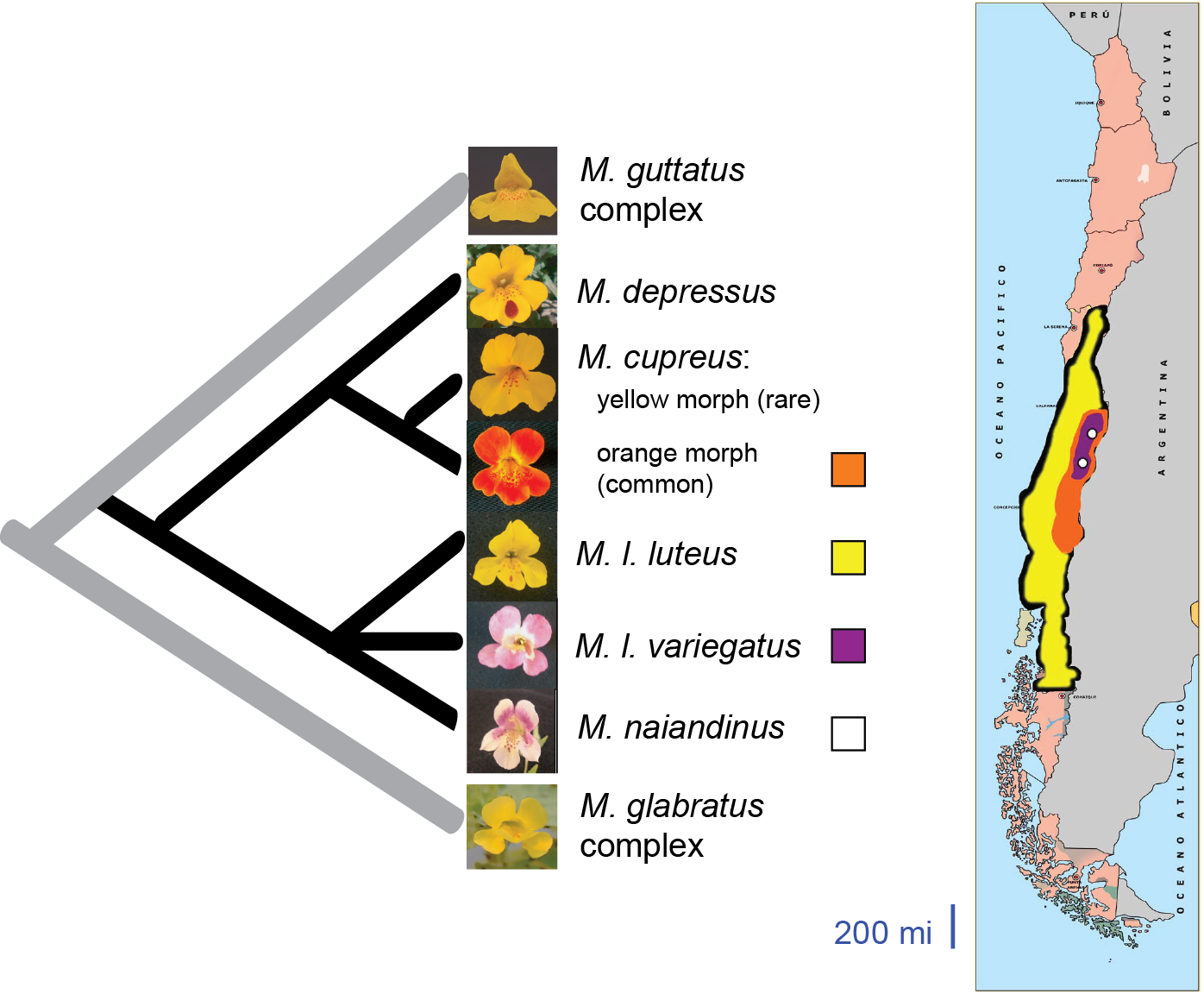
Related monkeyflower species, including genomic model M. guttatus, have a constant pigmentation pattern consisting of a yellow flower with small red spots of anthocyanin. In the Chilean Mimulus, in contrast, three taxa (M. cupreus, M. l. variegatus and M. naiandinus) have extensive anthocyanin pigment in their petals.
All three of these taxa produce yellow carotenoid pigment, plus a single type of red anthocyanin pigment called cyanidin. How can just two pigments produce such different colors? Click here to find out!
Dr. Cooley's prior work shows that M. l. variegatus and M. cupreus each recently gained extensive petal anthocyanin pigmentation, and in each case the pigment gain is controlled by a single genetic locus containing a cluster of anthocyanin-related MYB transcription factors. However, the pigmentation locus in M. l. variegatus is distinct from the M. cupreus locus. The two genomic regions are located on separate chromosomes and appear to be related by an ancestral gene or genome duplication event.
A major focus in the Cooley lab is to investigate the genetic and molecular basis of this interesting case of the parallel evoution of petal coloration. We will be using whole-genome data to identify and characterize all of the many recently-duplicated copies of the MYB transcription factors that seem to be rampant in the Chilean Mimulus genomes. This will be followed by qPCR expression studies and transgenic tests of gene function, to test each candidate MYB gene for a possible role in flower pigmentation.
Intriguing preliminary data suggest that M. naiandinus may represent a third, separate instance of the gain of petal anthocyanins. Did M. naiandinus also gain its anthocyanins thanks to evolution of a MYB transcription factor? Did one of the dozens of other anthocyanin-related genes play a role? Or perhaps we are wrong and M. naindinus did not "invent" its own red pigmentation at all - the range of M. naiandinus overlaps with that of both M. l. variegatus and M. cupreus, so it is possible that gene flow between species brought red petals either to or from M. naiandinus.
Finally, the anthocyanin biosynthetic pathway that is responsible for producing floral anthocyanins has a number of important physiological effects that I will be pursuing in the future - for example, they promote plant tolerance to heat, drought, and UV radiation. Is the parallel gain of petal anthocyanins, in M. cupreus and M. l. variegatus, associated with any changes in fitness relative to their less-heavily pigmented counterparts?